http://accessmedicine.mhmedical.com/CaseContent.aspx?gbosID=219724&gbosContainerID=65#105331831
Objectives
Describe the signaling mechanisms used in the nervous system.
Describe the necessary elements to create an electrochemical gradient.
Describe how the membrane and channels work to create the environment necessary for signaling.
Definitions
ELECTROCHEMICAL GRADIENT: The gradient across a cell membrane created by the differential concentrations of charged ions on either side of the membrane (for example, the difference in concentrations of potassium and sodium ions inside and outside a neuron that when regulated by their respective ion channels allow for signaling down an axon to occur).
ION CHANNELS: Transmembrane proteins in the cell membrane which open and close to allow for the passage of ions.
GATING: The process by which channels undergo conformational changes to allow for the passage of ions.
RESTING MEMBRANE POTENTIAL: The potential created in a neuron at rest by resting channels and sodium-potassium pumps.
Discussion
The nervous system depends on two types of signaling mechanisms, electrical and chemical, to propagate information throughout the nervous system. Rapid changes in the electrical potential across the neuronal cell membrane generate electrical signals that are transmitted down the length of the neuron. This system requires (1) an intact membrane to separate ions and maintain an electrochemical gradient and (2) ion channels to allow for the selective passage of ions of specific charges to generate the electrical signal.
The cell membrane of the neuron is formed by a lipid bilayer and is generally impermeable to charged particles. The double layer of phospholipids is hydrophobic. Charged ions are hydrophilic and as a result attract water molecules. This allows the neuronal cell membrane to separate charges across its surface to maintain the electrochemical gradient. However, to create and use the energy stored in the electrochemical gradient, structures must exist to allow for the passage of ions across this membrane. Ion channels, formed by transmembrane spanning proteins, serve that specific function within the neuron. The basic structure consists of transmembrane proteins with carbohydrate groups attached to their surface and a central pore-forming region to allow for the passage of ions. This pore-forming region spans the entirety of the membrane and is generally made up of two or more subunits.
Ion channels must also be selective for specific charged particles. One method by which channels select for specific ions is by size. Although the diameter of a potassium ion (K+) is larger than the diameter of a sodium ion (Na+), the Na+ ions demonstrate a stronger electrostatic attraction for water molecules. Thus, in a solution the Na+ ion has a larger shell of water than K+ ions. Channels can therefore select for K+ ions based upon the size differential in a solution. Other types of channels are selective for specific ions based upon the ion’s electrical affinity to charged portions of the channel. The attraction between an ion and the channel must be sufficiently strong enough to overcome the hydrostatic attraction of the ion. Once the shell of water surrounding the ion is shed, the ion can diffuse through the channel.
The flow of ions through a channel is passive and governed by the electrochemical gradient. Some ion channels are highly selective for a specific anion or cation, while others are more indiscriminate. Ion channels also open and close based upon the needs of the neuron. This change in state requires a conformational change of the proteins that form the channel, a process called gating.
To understand the electrical properties of the neuron, we must have an understanding of the electrochemical gradient. Particular ions are distributed unequally across the cell membrane. Concentrations of Na+ and Cl− are greater on the outside of the cell, while concentrations of K+ and organic anions, such as charged amino acids and proteins, are greater on the inside of the cell.
The figure below shows the mechanisms involved in the resting membrane potential across a cell membrane. There are 2 forces involved -
1) diffusion down concentration gradients
2) diffusion down electrical gradients
At equilibrium, these two forces are equal and opposite. The example given is for potassium, K+
At equilibrium, the chemical work = the electrical work. This equality allows you to rearrange the two equations to solve for membrane potential (Vm). At 37 C, RT/F = -60 mv.
 using normal values from table above, for K+
Vm = -60 x log 140/4 = -60 x log35 = -60 x 1.55 = -93 mv
using normal values from table above, for K+
Vm = -60 x log 140/4 = -60 x log35 = -60 x 1.55 = -93 mv
with hyperkalemia; e.g., Vm = -60 x log 140/6 = -60 x 1.36 = -81.6 mv
with hypokalemia; e.g., Vm = -60 x log 140/2
= -60 x 1.84 = -110 mv
Goldman Hodgkin Katz Equation
 Example with numbers
Example with numbers


In terms of myocardium cell potential:
hyperkalemia = depolarized resting potential, but Decreased excitability.
hypokalemia = hyperpolarized resting potential, but Increased excitability
In terms of EKG surface potential:
hyperkalemia = shortened QT, peaked T wave, wide QRS, ST depression
hypokalemia = prolonged QT, flat Twaves, U waves
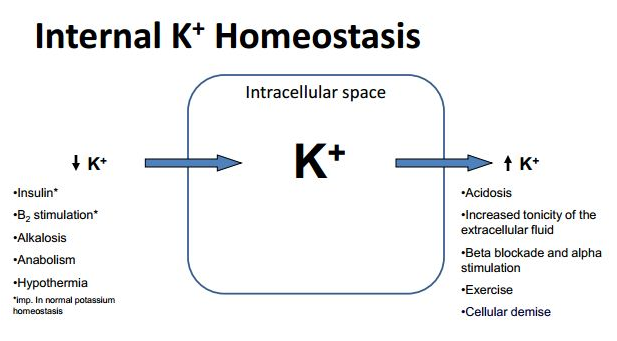
Potassium Homeostasis
Low serum potassium not only hyperpolarizes most cells, leading to an increase in the resting potential, but also has effects on certain potassium channels required for repolarization. Thus, hypokalemia decreases or slows potassium conductance, the prolonged repolarization phase accounting for the characteristic electrocardiographic findings of broad, flattened T waves. U waves are also indicative of this delay in repolarization . The patient is prone to tachyarrhythmias, including ventricular tachycardia.
Partial depolarization produces weaker action potentials and slower spread across myocardium
Inc. K+ = decreased size of Na+ channels, closure of inactivation gate, slow flux, slower conduction in myocardium; increased permeability to potassium. This decreases heart rate by the same mechanism as acetylcholine, reducing the slope of phase 4 of the sinoartrial node action potential.
- Depolarization decreases excitability because the rate of rise of phase 0 of the action potential (Vmax) is directly proportional to the value of the resting membrane potential at the onset of phase 0. This is because the membrane potential at the onset of depolarization determines the number of sodium channels activated during depolarization, which in turn determines the magnitude of the inward sodium current and the Vmax of the action potential.
 As illustrated in , Vmax is greatest when the resting membrane potential at the onset of the action potential is approximately −75 mV, and does not increase as the membrane potential becomes more negative. Conversely, as the resting membrane potential becomes less negative (that is, −70 mV), as in the setting of hyperkalemia (), the percentage of available sodium channels decreases. This decrease leads to a decrement in the inward sodium current and a concurrent decrease in the Vmax; therefore, as the resting membrane potential becomes less negative in hyperkalemia, Vmax decreases. This decrease in Vmax causes a slow-ing of impulse conduction through the myocardium and a prolongation of membrane depolarization; as a result, the QRS duration is prolonged.
As illustrated in , Vmax is greatest when the resting membrane potential at the onset of the action potential is approximately −75 mV, and does not increase as the membrane potential becomes more negative. Conversely, as the resting membrane potential becomes less negative (that is, −70 mV), as in the setting of hyperkalemia (), the percentage of available sodium channels decreases. This decrease leads to a decrement in the inward sodium current and a concurrent decrease in the Vmax; therefore, as the resting membrane potential becomes less negative in hyperkalemia, Vmax decreases. This decrease in Vmax causes a slow-ing of impulse conduction through the myocardium and a prolongation of membrane depolarization; as a result, the QRS duration is prolonged.
 Hydrogen enters cells in exchange for potassium
Hydrogen enters cells in exchange for potassium
 This does not occur in respiratory acidosis since CO2 is very lipid soluble and enters cell to produce H+ inside cell. No exchange for K+
This does not occur in respiratory acidosis since CO2 is very lipid soluble and enters cell to produce H+ inside cell. No exchange for K+

Excess H+ ions interfere with slow Ca++ channels (phase 2) and with binding to troponin C = decreased cardiac contractility
Net effect = muscle weakness, paralysis, cardiac arrhythmias
Hypokalemia
Potassium is the most abundant monovalent cation in body. concentration in intracellular space is about the same as that of Na in extracellular space (140 mEq/L). but the ICF volume is 2x ECF volume so there is twice as much K.
Em sensitive to extracellular K due to relatively small concentrations of K. Em not sensitive to same magnitude of change in intracellular fluid.
ECF has a total of 70 mEq of K. A meal may contain 70 mEq K. Portal vein receives K load, insulin is released, and insulin moves K into cells where the total K is 3500 mEq K. Then K slowly leaches out into blood to be excreted in urine.
“Ins and Outs” of K
NaK ATPase constantly bailing Na to keep K inside
K "bath" used to stop hearts for surgery
diarrhea loss can be much higher than nl (up to 100 mEq/day)
Muscle Weakness from hypokalemia
Hypokalemia will decrease potassium channel conductance, which will lengthen repolarization time of a nerve cell. If this gets to be severe enough, transmission of action potentials will be disrupted, and the result can be generalized weakness or paralysis because signaling to the muscles are disrupted. However, another possibility is cramping, the opposite effect, because of the same reason (being stuck in a depolarized state would also prevent the muscle from relaxing after a contraction was initiated).
In addition, the reduced extracellular potassium (paradoxically) inhibits the activity of the IKr potassium current[11] and delays ventricular repolarization. This delayed repolarization may promote reentrant arrythmias.
K Homeostasis
Kidneys reabsorb all filtered load of K (720 mEq/day) - most in collecting duct (principal cells)
Bartter's Syndrome - thick ascending limb = same effect as loop diuretics
Gittleman Syndrome - DCT - resembles thiazide diuretic
Potassium sparing diuretics